Halophiles are organisms that thrive in high salt concentrations. They are a type of extremophile organism. The name comes from the Greek word for "salt-loving". While most halophiles are classified into the Archaea domain, there are also bacterial halophiles and some eukaryota, such as the alga Dunaliella salina or fungus Wallemia ichthyophaga. Some well-known species give off a red color from carotenoid compounds, notably bacteriorhodopsin. Halophiles can be found anywhere with a concentration of salt five times greater than the salt concentration of the ocean, such as the Great Salt Lake in Utah, Owens Lake in California, the Dead Sea, and in evaporation ponds.
Classification[edit]
Halophiles are categorized as slight, moderate, or extreme, by the extent of their halotolerance. Slight halophiles prefer 0.3 to 0.8 M (1.7 to 4.8%—seawater is 0.6 M or 3.5%), moderate halophiles 0.8 to 3.4 M (4.7 to 20%), and extreme halophiles 3.4 to 5.1 M (20 to 30%) salt content.[1] Halophiles require sodium chloride (salt) for growth, in contrast to halotolerant organisms, which do not require salt but can grow under saline conditions.
Lifestyle[edit]
High salinity represents an extreme environment to which relatively few organisms have been able to adapt and occupy. Most halophilic and all halotolerant organisms expend energy to exclude salt from their cytoplasm to avoid protein aggregation ('salting out'). To survive the high salinities, halophiles employ two differing strategies to prevent desiccationthrough osmotic movement of water out of their cytoplasm. Both strategies work by increasing the internal osmolarity of the cell. In the first (which is employed by the majority of halophilic bacteria, some archaea, yeasts, algae and fungi), organic compounds are accumulated in the cytoplasm—osmoprotectants which are known as compatible solutes. These can be either synthesised or accumulated from the environment.[2] The most common compatible solutes are neutral or zwitterionic, and include amino acids, sugars, polyols, betaines, and ectoines, as well as derivatives of some of these compounds.
The second, more radical adaptation involves the selective influx of potassium (K+) ions into the cytoplasm. This adaptation is restricted to the moderately halophilic bacterial order Halanaerobiales, the extremely halophilic archaeal family Halobacteriaceae, and the extremely halophilic bacterium Salinibacter ruber. The presence of this adaptation in three distinct evolutionary lineages suggests convergent evolution of this strategy, it being unlikely to be an ancient characteristic retained in only scattered groups or passed on through massive lateral gene transfer.[2] The primary reason for this is the entire intracellular machinery (enzymes, structural proteins, etc.) must be adapted to high salt levels, whereas in the compatible solute adaptation, little or no adjustment is required to intracellular macromolecules; in fact, the compatible solutes often act as more general stress protectants, as well as just osmoprotectants.[2]
Of particular note are the extreme halophiles or haloarchaea (often known as halobacteria), a group of archaea, which require at least a 2 M salt concentration and are usually found in saturated solutions (about 36% w/v salts). These are the primary inhabitants of salt lakes, inland seas, and evaporating ponds of seawater, such as the deep salterns, where they tint the water column and sediments bright colors. These species most likely perish if they are exposed to anything other than a very high-concentration, salt-conditioned environment. These prokaryotes require salt for growth. The high concentration of sodium chloride in their environment limits the availability of oxygen for respiration. Their cellular machinery is adapted to high salt concentrations by having charged amino acids on their surfaces, allowing the retention of water molecules around these components. They are heterotrophs that normally respire by aerobic means. Most halophiles are unable to survive outside their high-salt native environments. Indeed, many cells are so fragile that when placed in distilled water, they immediately lyse from the change in osmotic conditions.
Halophiles may use a variety of energy sources. They can be aerobic or anaerobic. Anaerobic halophiles include phototrophic, fermentative, sulfate-reducing, homoacetogenic, and methanogenic species.[1][3]
The Haloarchaea, and particularly the family Halobacteriaceae, are members of the domain Archaea, and comprise the majority of the prokaryotic population in hypersaline environments.[4] Currently, 15 recognised genera are in the family.[5] The domain Bacteria (mainly Salinibacter ruber) can comprise up to 25% of the prokaryotic community, but is more commonly a much lower percentage of the overall population.[6] At times, the alga Dunaliella salina can also proliferate in this environment.[7]
A comparatively wide range of taxa has been isolated from saltern crystalliser ponds, including members of these genera: Haloferax, Halogeometricum, Halococcus, Haloterrigena, Halorubrum, Haloarcula, and Halobacterium.[4] However, the viable counts in these cultivation studies have been small when compared to total counts, and the numerical significance of these isolates has been unclear. Only recently has it become possible to determine the identities and relative abundances of organisms in natural populations, typically using PCR-based strategies that target 16S small subunit ribosomal ribonucleic acid (16S rRNA) genes. While comparatively few studies of this type have been performed, results from these suggest that some of the most readily isolated and studied genera may not in fact be significant in the in situ community. This is seen in cases such as the genus Haloarcula, which is estimated to make up less than 0.1% of the in situ community,[8] but commonly appears in isolation studies.
Genomic and proteomic signature[edit]
The comparative genomic and proteomic analysis showed distinct molecular signatures exist for environmental adaptation of halophiles. At the protein level, the halophilic species are characterized by low hydrophobicity, overrepresentation of acidic residues, underrepresentation of Cys, lower propensities for helix formation, and higher propensities for coil structure. The core of these proteins is less hydrophobic, such as DHFR, that was found to have narrower β-strands.[9] At the DNA level, the halophiles exhibit distinct dinucleotide and codon usage.[10]
Examples[edit]
Halobacterium[11] is a genus of the Archaea that has a high tolerance for elevated levels of salinity. Some species of halobacteria have acidic proteins that resist the denaturing effects of salts. Halococcus is a specific genus of the family Halobacteriaceae.
Some hypersaline lakes are a habitat to numerous families of halophiles. For example, the Makgadikgadi Pans in Botswana form a vast, seasonal, high-salinity water body that manifests halophilic species within the diatom genus Nitzschia in the family Bacillariaceae, as well as species within the genus Lovenula in the family Diaptomidae.[12] Owens Lake in California also contains a large population of the halophilic bacterium Halobacterium halobium.
Wallemia ichthyophaga is a basidiomycetous fungus, which requires at least 1.5 M sodium chloride for in vitro growth, and it thrives even in media saturated with salt.[13] Obligate requirement for salt is an exception in fungi. Even species that can tolerate salt concentrations close to saturation (for example Hortaea werneckii) in almost all cases grow well in standard microbiological media without the addition of salt.[14]
The fermentation of salty foods (such as soy sauce, Chinese fermented beans, salted cod, salted anchovies, sauerkraut, etc.) often involves halobacteria, as either essential ingredients or accidental contaminants. One example is Chromohalobacter beijerinckii, found in salted beans preserved in brine and in salted herring. Tetragenococcus halophilus is found in salted anchovies and soy sauce.
Artemia is a ubiquitous genus of small halophilic crustaceans living in salt lakes (such as Great Salt Lake) and solar salterns that can exist in water approaching the precipitation point of NaCl, 340 g L−1[15][16] and can withstand strong osmotic shocks thanks to its mitigating strategies for fluctuating salinity levels, such as its unique larval salt gland and osmoregulatory capacity.
North Ronaldsay sheep are a breed of sheep originating from Orkney, Scotland. They have limited access to fresh water sources on the island and to their only food source is seaweed. They have adapted to handle salt concentrations that would kill other breeds of sheep.[17]
Halotolerance is the adaptation of living organisms to conditions of high salinity.[1] Halotolerant species tend to live in areas such as hypersaline lakes, coastal dunes, saline deserts, salt marshes, and inland salt seas and springs. Halophiles are organisms that live in highly saline environments, and require the salinity to survive, while halotolerant organisms (belonging to different domains of life) can grow under saline conditions, but do not require elevated concentrations of salt for growth. Halophytes are salt-tolerant higher plants. Halotolerant microorganisms are of considerable biotechnological interest.[2]
Applications[edit]
Fields of scientific research relevant to halotolerance include biochemistry, molecular biology, cell biology, physiology, ecology, and genetics.
An understanding of halotolerance can be applicable to areas such as arid-zone agriculture, xeriscaping, aquaculture (of fish or algae), bioproduction of desirable compounds (such as phycobiliproteins or carotenoids) using seawater to support growth, or remediation of salt-affected soils. In addition, many environmental stressors involve or induce osmotic changes, so knowledge gained about halotolerance can also be relevant to understanding tolerance to extremes in moisture or temperature.
Goals of studying halotolerance include increasing the agricultural productivity of lands affected by soil salination or where only saline water is available. Conventional agricultural species could be made more halotolerant by gene transfer from naturally halotolerant species (by conventional breeding or genetic engineering) or by applying treatments developed from an understanding of the mechanisms of halotolerance. In addition, naturally halotolerant plants or microorganisms could be developed into useful agricultural crops or fermentation organisms.
Cellular functions in halophytes[edit]
Tolerance of high salt conditions can be obtained through several routes. High levels of salt entering the plant can trigger ionic imbalances which cause complications in respiration and photosynthesis, leading to reduced rates of growth, injury and death in severe cases. To be considered tolerant of saline conditions, the protoplast must show methods of balancing the toxic and osmotic effects of the increased salt concentrations. Halophytic vascular plants can survive on soils with salt concentrations around 6%, or up to 20% in extreme cases. Tolerance of such conditions is reached through the use of stress proteins and compatible cytoplasm osmotic solutes.[3]
To exist in such conditions, halophytes tend to be subject to the uptake of high levels of salt into their cells, and this is often required to maintain an osmotic potential lower than that of the soil to ensure water uptake. High salt concentrations within the cell can be damaging to sensitive organelles such as the chloroplast, so sequestration of salt is seen. Under this action, salt is stored within the vacuole to protect such delicate areas. If high salt concentrations are seen within the vacuole, a high concentration gradient will be established between the vacuole and the cytoplasm, leading to high levels of energy investment to maintain this state. Therefore, the accumulation of compatible cytoplasmic osmotic solutes can be seen to prevent this situation from occurring. Amino acids such as proline accumulate in halophytic Brassica species, quaternary ammonium bases such as Glycine Betaine and sugars have been shown to act in this role within halophytic members of Chenopodiaceae and members of Asteraceae show the buildup of cyclites and soluble sugars. The buildup of these compounds allow for the balancing of the osmotic effect while preventing the establishment of toxic concentrations of salt or requiring the maintenance of high concentration gradients
Bacterial halotolerance[edit]
The extent of halotolerance varies widely amongst different species of bacteria.[4] A number of cyanobacteria are halotolerant; an example location of occurrence for such cyanobacteria is in the Makgadikgadi Pans, a large hypersaline lake in Botswana.[5]
Fungal halotolerance[edit]
Fungi from habitats with high concentration of salt are mostly halotolerant (i.e. they do not require salt for growth) and not halophilic. Halophilic fungi are a rare exception.[6]Halotolerant fungi constitute a relatively large and constant part of hypersaline environment communities, such as those in the solar salterns.[7] Well studied examples include the yeast Debaryomyces hansenii and black yeasts Aureobasidium pullulans and Hortaea werneckii.[8] The latter can grow in media without salt, as well as in almost saturated NaClsolutions. To emphasize this unusually wide adaptability, some authors describe H. werneckii as "extremely halotolerant".[9]
The world of halophilic microorganisms is highly diverse. Microbes adapted to life at high salt concentrations are found in all three domains of life: Archaea, Bacteria, and Eucarya. In some ecosystems salt-loving microorganisms live in such large numbers that their presence can be recognized without the need for a microscope. The brines of saltern crystallizer ponds worldwide are colored pink-red by Archaea (Haloquadratum and other representatives of the Halobacteriales), Bacteria (Salinibacter), and Eucarya (Dunaliella salina).
Hypersaline environments such as saltern pond brines and natural salt lakes present the ecologist with relatively simple ecosystems with low diversity and high community densities. In such systems fundamental questions of biodiversity, selection, biogeography, and evolution in the microbial world can be investigated much more conveniently than in the far more complex freshwater and marine systems. The sediments of such water bodies, however, are often inhabited by extremely diverse, still incompletely explored microbial communities. Different types of halophiles have solved the problem how to cope with salt stress (and often with other forms of stress as well) in different ways, so that the study of microbial life at high salt concentrations can answer many basic questions on the adaptation of microorganisms to their environments. Most known halophiles are relatively easy to grow, and genera such as Halobacterium, Haloferax, and Haloarcula have become popular models for studies of the archaeal domain as they are much simpler to handle than methanogenic and hyperthermophilic Archaea. Some halophilic and halotolerant microorganisms have found interesting biotechnological applications as well, as shown in the last section of this report.
The 9th International Conference on Halophilic Microorganisms, held from 29 June 2010 to 3 July 2010 in Beijing, China, brought together 166 participants from 25 countries. The 50 lectures and 112 posters presented provided an excellent overview of the current state of our understanding of all aspects of microbiology at high salt concentrations. The meeting was hosted by the Institute of Microbiology, the Chinese Academy of Sciences, the Chinese Society of Biotechnology, and the Chinese Society for Microbiology. Conference chair was Yanhe Ma. The series of symposia on halophiles started in Rehovot, Israel, in 1978 with a meeting devoted mainly to the properties of bacteriorhodopsin, the retinal-containing protein of Halobacteriumthat was discovered just a few years earlier. The delegates noted and applauded the presence of Janos Lanyi in the audience, one of the attendees at the first meeting. This initial event was followed by meetings held in 1985 (Obermarchtal, Germany), 1989 (Alicante, Spain), 1992 (Williamsburg, VA), 1997 (Jerusalem, Israel), 2001 (Seville, Spain), 2004 (Ljubljana, Slovenia), and 2007 (Colchester, United Kingdom). The proceedings of the 1978, 1989, 1997, 2001, and 2004 symposia were published as books (20, 31, 56, 68, 88); selected papers from the 1985, 2002, and 2007 symposia appeared in dedicated special volumes of journals (FEMS Microbiology Reviews, Experientia, and Saline Systems).
This review intends to capture emerging themes and to report key interesting new findings presented at the Halophiles 2010 symposium in Beijing. The following topics were the focus of attention and discussion.
DIVERSITY OF HALOPHILES—CULTURED AND UNCULTURED
“Everything is everywhere: but, the environment selects” (“Alles is overal: maar, het milieu selecteert”). This famous quotation from Lourens Baas Becking's 1934 book Geobiologie of inleiding tot the milieukunde (8) can be taken as the basis for our understanding of the distribution of halophilic microorganisms worldwide. In fact, Baas Becking (1895 to 1963) had visited many salt lakes and studied many different halophilic microorganisms. His book and his publications from the early 1930s contain a wealth of information, largely forgotten today, on the properties of the halophiles. Some phenomena described by Baas Becking at the time, including the acidic nature of the cell envelope of Dunaliella and the interrelationship of salt requirement/tolerance and temperature in halophilic prokaryotes, were “rediscovered” in the 1970s, as documented by Aharon Oren (Jerusalem, Israel) in his keynote lecture at the opening session.
To explore to what extent in the halophilic world “everything” is indeed “everywhere” and what degree of variation may be found among different high-salt environments, microbial diversity studies have been performed in a great variety of environments. These include saltern ponds worldwide, Great Salt Lake, the Dead Sea (16), saline lakes in Inner Mongolia (60), African soda lakes, deep-sea brines (85), and many others. These studies included culture-dependent approaches, leading to the isolation and characterization of many novel types of halophiles and new information on the abundance and geographic distribution of the known types, as well as culture-independent studies based on sequencing of DNA recovered from the environment. Many posters at the meeting related to culture-independent analyses of hypersaline environments from around the world, including Xinjiang salt lakes; Chinese salt mines; salterns in Goa India, Turkey, Spain, and Israel; south Siberian hypersaline lakes; the Dead Sea; and Great Salt Lake. Thane Papke (Storrs, CT) examined Halorubrum strains in Spain and Algeria, and one of his conclusions was that “migration routes are slower than mutation rates,” allowing endemism in Halorubrum strains to develop. Shaun Heaphy (Leicester, United Kingdom) provided a culture-independent microbial characterization of several Inner Mongolian salt and soda lakes and used statistical techniques to correlate the findings with physico-chemical parameters of brines and geographical location. He broadly agreed that microbial populations diverged as distance between the lakes increased, although this was only statistically significant for the Bacteria on an intercontinental scale (a hypersaline lake in Argentina was included in the analysis). Factors such as pH, temperature, and Na+ concentration were particularly correlated with the microbial community composition. Thus, everything may not be everywhere. More and more cases are being reported of the isolation of halophilic microorganisms from low salinity environments. Thus, after almost 80 years, Baas Becking's quotation still inspires experiment and debate.
Application of culture-dependent methods led to the isolation of a novel halophilic archaeon from seawater (at a salinity that does not support growth of Halobacteriaceae and causes lysis of most known representatives of the group). The properties of this new organism, to be described as a new genus and species, Halomarina oriensis, were presented by Kentaro Inoue (Chiba, Japan) (37). Poster presentations included a new fungal isolate from a Turkish salt mine; novel haloarchaea from a Chinese saltern, Inner Mongolian lakes, and Iranian salt lakes; and novel bacterial isolates from Chinese salt lakes, Inner Mongolian Lakes, Xinjiang salt lakes, Quidam Basin Quaternary sediments, brine wells in southwestern China, the Yellow Sea, The South China Sea, a Korean salt flat, Iranian salt lakes, Mexican soda environments, and salted hides. There were additional reports on the isolation of new actinomycetes from saline environments in China and on different bacterial halophiles from nonsaline sites such as soils.
HALOPHILES IN UNUSUAL ENVIRONMENTS AND HALOPHILES EXPOSED TO MULTIPLE FORMS OF STRESS
Most habitats explored for the presence of halophiles are thalassohaline environments that originated by evaporation from seawater, reflect the ionic composition of seawater, and have a nearly neutral to slightly alkaline pH.
Deep-sea brines, found on the bottom of the Red Sea, the Mediterranean Sea, and the Gulf of Mexico, are interesting environments to search for novel microbes. Apart from their increased high salinity, they are anaerobic and form characteristically sharp brine-seawater interfaces, with some of the brines displaying significant increases in temperature and metal concentration. The ionic composition of the brines generally differs from that of seawater; they are anaerobic, and in some cases the temperature can be elevated as well. The microbiology of Shaban Deep and other deep-sea brines in the Red Sea was discussed by André Antunes (Thuwal, Saudi Arabia). These sites, considered sterile in the past, have yielded a number of interesting microorganisms, including Salinisphaera shabanensis (a facultative anaerobe growing in a very large range of salt concentrations, from 1 to 28%) (5), Halorhabdus tiamatea (a nonpigmented representative of the Halobacteriales that prefers an anaerobic life style) (7), Flexistipes sinusarabici (an anaerobe tolerating between 3 to 18% NaCl) (28), and Haloplasma contractile(a contractile bacterium, phylogenetically equidistant to the Firmicutes and the Mollicutes) (6). The sites will be revisited in the near future for further microbiological exploration.
In many athalassohaline environments, life at the extremes of high salt is combined with the need to thrive at alkaline pH and elevated temperatures, and organisms growing there do so at the physico-chemical boundary for life (18). Jürgen Wiegel (Athens, GA) summarized his studies of the anaerobic halophilic, alkaliphilic, thermophilic bacteria isolated from the Wadi an-Natrun, Egypt. Natranaerobius thermophilus accumulates both glycine betaine and K+ for osmotic adaptation and has multiple Na+/H+ antiporters (49) and a Na+-extruding ATPase, which was characterized in-depth by Noha Mesbah (Alexandria, Egypt). Two new species, designated “Natranaerobius jonesii” and “Natranaerobius grantii” are currently being characterized. Natranaerobius jonesii has an extremely high requirement for chloride ions as it does not grow at less than 1.4 M Cl−. Other alkaline saline environments subjected to intensive studies in recent years are the soda lakes of the Kulunda Steppe (Altai, Russia). Dimitry Sorokin (Moscow, Russia) summarized the wealth of information obtained from these studies at the level of the characterization of cultures of novel organisms, especially those participating in the reductive part of the sulfur cycle, and from culture-independent studies using molecular markers, as well as measurements of the rates of microbial sulfidogenesis. In general, sulfide production was active even in saturated soda brines, but far more sulfide was produced in these environments from elemental sulfur and from thiosulfate than from sulfate. Dismutation of thiosulfate and sulfite was a major trend in soda lake isolates (78, 79).
The Dead Sea is a rare example of a low-Na+, high-Mg2+, and high-Ca2+ chloride brine with a slightly acidic pH. Metagenomic studies are now providing information on the microbial diversity in the lake, both at the time of a bloom of microorganisms following dilution of the upper water layers by rain floods in 1992 and during the current drying out of the lake, causing a continuously decreasing ratio of monovalent/divalent cations, making conditions too extreme for even the best salt-adapted microorganisms (16).
GENOMICS, EVOLUTION, AND TAXONOMY OF HALOPHILIC ARCHAEA AND BACTERIA
Genomics of the Halobacteriaceae has come of age. Shiladitya DasSarma (Baltimore, MD) gave the closing keynote lecture, highlighting the haloarchaeal genomes from different genera which have been determined since the genome sequence of Halobacterium NRC-1 was first published (24, 53) 10 years ago. More recently completed genomes highlighted included Haloarcula marismortui (11, 14), Natronomonas pharaonis (26), Haloquadratum walsbyi (17), Halorubrum lacusprofundi (http://www.ncbi.nlm.nih.gov/sites/entrez?Db=genome&Cmd=ShowDetailView&TermToSearch=23834, 2009), Halomicrobium mukohataei(82), Halorhabdus utahensis (10), Halogeometricum borinquense (46), Haloterrigena turkmenica (73), and Haloferax volcanii (34). The list includes significant ecological diversity, e.g., a haloalkaliphilic species, a cold-adapted species, species adapted to life in low-Na+-high-Mg2+ environments, and isolates showing interesting cell morphologies. The sizes of these genomes range between 2.6 and 5.4 Mb. The sequencing and analysis of the genomes of Haloarcula hispanica and Haloferax mediterranei were announced by Hua Xiang and colleagues (Beijing, China). DasSarma showed that there has been an exponential increase in the sequencing of haloarchaeal genomes over the past 10 years, and, with next-generation sequencing methods now available, expects that within a few years the number of published genomes of species of Halobacteriaceae will grow even faster. Some of the conserved properties of haloarchaeal genomes were discussed, including the presence of large megaplasmids and minichromosomes (24) and the occurrence of core acidic proteomes (23). The data analysis also yielded the prediction of an expanding haloarchaeal pan-genome with increasing numbers of novel genes which may have applications in biotechnology.
In two presentations from the DasSarma group, additional postgenomic work was presented. James Coker (soon to move to Birmingham, AL) reported on studies on the expanded TATA-binding protein and transcription factor B protein families of haloarchaea, showing their importance for gene expression and stress regulation (21, 77). Satyajit DasSarma (the youngest presenter at age 13) reported on the expansion of the HaloWeb, the haloarchaeal genome database (http://halo4.umbi.umd.edu), which now provides access to all the public haloarchaeal genomes and as well as a suite of tools for data retrieval and analysis.
Environmental genomics studies increasingly show that the genome of individual strains may be only a small fraction of the pan-genome of the species in nature. Haloquadratum walsbyihas become an excellent example to illustrate this, as shown by Francisco Rodríguez-Valera (Alicante, Spain) and Mike Dyall-Smith (Martinsried, Germany). Comparisons have been made of the genome diversity within Haloquadratum populations in a single saltern crystallizer pond as well as comparisons between populations in similar environments at different geographic locations. The pan-genome of Haloquadratum walsbyi is at least 40 times the size of the genome of the type strain, and genomic microdiversity within an extremely simple and relatively constant environment is very high (17, 22, 40, 55). An interesting study of experimental evolution was presented by Jizhong Zhou (Norman, OK), using Desulfovibrio vulgaris as a model organism and monitoring genetic changes after exposure of this nonhalophilic bacterium to 0.25 M NaCl for 1,000 generations. Salt-specific mutations and deletions were detected in the salt-resistant phenotype, which used different amino acids as osmoprotectants (35).
Genome sequencing of new isolates is getting simpler and cheaper and will probably soon become routine. Undoubtedly, this development will have profound implications on the taxonomy of the halophiles. Until taxonomy can be based on comparison of complete genome sequences, multilocus sequence analysis (MLSA) is gaining popularity for the comparison of strains for taxonomic and evolutionary studies. Thane Papke (Storrs, CT) presented his extensive MLSA data on Halorubrum isolates from Spain and Algeria. Analysis of the data indicates very frequent occurrence of homologous recombination, to the extent that alleles were randomly associated, as typical of sexually reproducing species. Natural competence and conjugation (like the mating mechanism in Haloferax) (3) may be the possible mechanisms for lateral gene transfer (61). Emma White (Storrs, CT) and Hiroaki Minegishi (Saitama, Japan) showed how analysis of the RNA polymerase subunit B′(rpoB′) gene can help in reconstructing the phylogeny of the Halobacteriaceae (51). Also for the Halomonadaceae, MLSA is becoming a valuable tool for taxonomic studies, as shown by Antonio Ventosa (Seville, Spain). For both groups, sets of genes and primers have been defined that give good results consistent with other genotypic and phenotypic traits.
The list of sequenced genomes of halophilic and halotolerant Bacteria is as yet short. It does not yet even include Halomonas elongata, the organism that, since it was described 30 years ago (89), has become one of the most popular model organisms and has also found biotechnological applications (30, 57). Its genome sequence will soon be published. Genome sequence information is available for the anoxygenic halophilic phototroph Halorhodospira halophila, for an extremely salt-tolerant alkaliphilic sulfur-oxidizing bacterium of the genus Thioalkalivibrio, for the thermophilic anaerobic halophile Halothermothrix orenii, and for the aerobic heterotrophic Chromohalobacter salexigens (58) and Salinibacter ruber.
Extensive environmental genomics data have been collected for Salinibacter. Josefa Antón (Alicante, Spain) showed a high degree of genomic variation within Salinibacter populations. Comparative analyses indicate that Salinibacter ruber genomes present a mosaic structure with conserved and hypervariable regions. Overall, 10% of the genes encoded in the genome of the Salinibacter M8 genome are absent from the type strain Salinibacter M31. Metabolomic profiles also differed in these two isolates (62).
HALOPHILIC VIRUSES
The Halophiles 2001 and 2004 symposia in Seville and Ljubljana will be remembered as the events where the importance of fungi in hypersaline ecosystems became clear. Halophiles 2010 can then be described as the congress presenting the importance of viruses. Phages attacking extremely halophilic Archaea were first described already in 1974 (83), but the role of viruses in hypersaline ecosystems remained largely unexplored.
Elina Roine (Helsinki, Finland) and her colleagues have discovered novel types of viruses attacking halophilic Archaea. The isolation and characterization of pleomorphic viruses possessing a lipid envelope, containing either a single-stranded or double-stranded DNA genome, show that viral diversity in hypersaline environments (63, 64, 69) is much larger than previously assumed. Shaun Heaphy (Leicester, United Kingdom) presented two novel lytic head/tailed viruses (virus BJ1 of the Siphoviridae and virus BJ2 of the Myoviridae), infecting Halorubrum kocurii, isolated from a salt lake in Inner Mongolia (59). Few archaeal virus genomes have been sequenced, and the complete sequence of virus BJ1 (EMBL accession number AM419438) is therefore a welcome addition.
First results of a comprehensive study of viral distribution and diversity in Great Salt Lake, UT, were presented by Bonnie Baxter (Salt Lake City, UT). Saltern crystallizer ponds are also ideal environments to study virus diversity and dynamics, as protozoa and other predators are absent, and numbers of prokaryotes and virus-like particles are extremely high, typically in the order of >107/ml and >108 to 109/ml, respectively. Forest Rohwer (San Diego, CA) showed his studies of virus dynamics in such salt-saturated ponds. At first sight, the salterns present predictable and stable communities of both Archaea and viruses, apparently different from the “kill-the-winner” behavior, with rapid cycling of microbial taxa and their viral predators that may be expected in such an environment (67). Metagenomic analysis of the viruses in the salterns near San Diego showed that the distribution of microbial taxa and viral taxa remained stable over time but with strong dynamic fluctuations of the prevalence of microbial strains and viral genotypes. Thus, at the fine level, the populations of both individual strains and viral genotypes fluctuate in a kill-the-winner fashion (67).
Activity of viruses also may have profound implications on the distribution of the extremely halophilic bacterium Salinibacter (Bacteroidetes). Josefa Antón (Alicante, Spain) studied the metagenome of viral assemblages of saltern pond in which Salinibacter accounts for around 15% of the prokaryotic community. Based on bioinformatic analysis (G+C content and dinucleotide frequency analysis), about 24% of the retrieved viral sequences could correspond to Salinibacter phages (70). It seems that phages infecting Salinibacter are more active in the environment than phages infecting Haloquadratum, and this may possibly explain why Haloquadratum outnumbers Salinibacter in every environment that supports growth of these organisms.
HALOPHILIC FUNGI
The importance of halophilic fungi, long neglected as members of hypersaline ecosystems, became recognized only in the past decade. Nina Gunde-Cimerman (Ljubljana, Slovenia) gave an overview of the biology of the most widespread and most halophilic or halotolerant fungi and yeasts. These include the black yeasts Hortaea werneckii which grows up to 5 M NaCl, the true halophile Wallemia ichthyophaga that requires at least 1.5 M NaCl and grows up to saturation, and Aureobasidium pullulans that grows up to 3 M NaCl. All of these are commonly found in hypersaline lakes and in a great variety of other, often unexpected, environments: domestic dishwashers, polar ice, and possibly even on spider webs in desert caves (32).
The halophilic and halotolerant fungi use polyols such as glycerol, erythritol, arabitol, and mannitol as osmotic solutes and retain low salt concentrations in their cytoplasm. Molecular studies on osmotic adaptation of Hortaea werneckii and Wallemia ichthyophaga were presented by Ana Plemenitaš and Janja Zajc (Ljubljana, Slovenia). Identification and structural features of Na+-sensitive 3′-phosphoadenosine-5′-phosphatase HwHal2, one of the putative determinants of halotolerance in H. werneckii and a promising transgene to improve halotolerance in crops, was presented (87). An in-depth understanding has been obtained of the HOG (high osmolarity glycerol) pathway, and this understanding may be applied in the future to the development of improved salt-resistant crops. Glycerol-3-phosphate dehydrogenase is involved in glycerol synthesis by both Wallemia and Hortaea, and heterologous expression of the gene encoding the enzyme can restore halotolerance in Saccharomyces cerevisiae deficient in glycerol production.
LONG-TERM SURVIVAL OF HALOPHILES
When brines dry out and halite crystals are formed, small fluid inclusions remain entrapped within the crystals. Microorganisms that inhabited the brine may get entrapped in these inclusions (9). Since the first controlled studies showed that such microorganisms may retain their viability for long periods (54), the question of the longevity of different types of halophiles within salt crystals has become a popular topic, relevant to disciplines including geology, biogeography, evolution, and even space exploration (48).
Terry McGenity (Colchester, United Kingdom) presented field studies and laboratory simulations of entombment of different types of microbes inside salt crystals. Salinibacter alone survives poorly within halite crystals, but when it was trapped inside a crystal together with Haloquadratum, longevity was much enhanced. Thus, simple food chains and mutual interactions occur between microorganisms in fluid inclusions in salt.
Examination of halite cores from Saline Valley, CA, representing salt deposited up to 150 thousand years ago, showed remnants of algae within fluid inclusions entrapped in the salt crystals. Morphological features as well as sequences of the internal transcribed spacer between the 18S and 5.8S rRNA genes led to the identification of Dunaliella, Ulothrix, and Nephroselmis, as shown by Krithivasan Sankaranarayanan (Binghamton, NY), who won first prize for his presentation by a young scientist. Presence of entrapped algae, with their high content of organic compatible solutes, may provide carbon and energy sources enabling halophilic heterotrophic microorganisms to survive for prolonged times (75).
OSMOTIC ADAPTATION, COMPATIBLE SOLUTES, AND ADAPTATION OF INTRACELLULAR PROTEINS TO SALT
There are basically two strategies that enable halophilic and halotolerant microorganisms to live in high salt concentrations. The “high-salt-in” strategy (used by the Halobacteriaceae, Salinibacter, and the anaerobic Halanaerobiales) requires all intracellular proteins to be stable and active in the presence of molar concentrations of KCl and other salts. The “low-salt, organic-solutes-in” strategy is based on the biosynthesis and/or accumulation of organic solutes that do not interfere greatly with the activity of normal enzymes. But even such organisms need to have salt-adapted proteins in the membrane exposed to the saline medium. It is remarkable that already in the early 1930s Baas Becking concluded that Dunaliella must have a highly acidic surface, based on the insensitivity of the alga to certain otherwise toxic anions (8).
Over the years, Haloarcula marismortui has been the most popular model organism for the study of the behavior of proteins active in a high-salt environment. These include the Haloarcula ribosome, whose structure elucidation by Ada Yonath was awarded the Nobel Prize for Chemistry in 2009. Christine Ebel (Grenoble, France) presented an overview of molecular adaptations of halophilic proteins, based on her studies of the Haloarcula marismortui malate dehydrogenase and other enzymes. Particularly, the very acidic surface of the macromolecule allows protein-salt interactions that avoid water or salt enrichment at the surface of the protein at high salt and preserve its solubility (25, 44).
Volker Müller (Frankfurt, Germany) uses Halobacillus halophilus as a model to understand the mechanisms of osmotic adaptation by a bacterium that accumulates organic-compatible solutes. Using techniques of biochemistry, genomics, DNA microarrays, etc., his group studies the way the organism senses its environment. Halobacillus is the first chloride-dependent bacterium reported, and several cellular functions depend on Cl− for maximal activities, the most important being the activation of solute accumulation. Halobacillus switches its osmolyte strategy with the salinity in its environment by the production of different compatible solutes. Glutamate and glutamine dominate at intermediate salinities, and proline and ectoine dominate at high salinities. Chloride stimulates expression of the glutamine synthetase and activates the enzyme. The product glutamate then turns on the biosynthesis of proline by inducing the expression of the proline biosynthetic genes (71, 72). Halobacillus dabanensis is used by Su-Sheng Yang and his colleagues (Beijing, China) as a model organism to study the genes involved in halotolerance, including genes encoding Na+/H+ antiporters, enzymes involved in osmotic solute metabolism, and stress proteins (27, 90). Studies of a mutant of Halomonas elongata deficient in ectoine synthesis by Elisabeth Witt (Bonn, Germany) showed the production of a new cyclic compatible solute, 5-amino-3,4-dihydro-2H-pyrrole-2-carboxylate (ADPC). It is made by a side reaction of ectoine synthase (EctC) that forms ADPC by cyclic condensation of glutamine. She also demonstrated that ectoine synthase is a reversible enzyme, which has its equilibrium (in case of ectoine synthesis) completely on the side of the cyclic condensation product.
Ectoine and hydroxyectoine biosynthesis is widely found in halophilic and halotolerant microorganisms, and the expression of the ect structural genes is induced by salt stress. But the solutes provide protection not only against salt stress but also against temperature stress in Bacillus subtilis and other salt-tolerant bacilli, as shown by Erhard Bremer (Marburg, Germany). Quantification of the intracellular ectoine concentration in Virgibacillus pantothenticus revealed that its production is triggered either by an increase in external salinity or by a reduction in growth temperature. Transcription of the ectoine biosynthetic operon (ectABC) was enhanced under both environmental conditions (38). The crystal structure of Virgibacillus salexigens EctD, the enzyme responsible for conversion of ectoine to hydroxyectoine, is now known in detail (66).
LIPIDS, MEMBRANE-BOUND PIGMENTS, AND MEMBRANE-LINKED PROCESSES
The cytoplasmic membranes of halophilic Archaea of the family Halobacteriaceae contain interesting ether lipids and often have retinal proteins (bacteriorhodopsin, halorhodopsin, and sensory rhodopsins). Interesting lipids and retinal proteins have also been found in Salinibacter.
Heiko Patzelt (Muscat, Oman) showed that unsaturated ether lipids are far more common in the halophilic Archaea than generally assumed. Such unsaturated diether lipids were earlier reported from the psychrotolerant haloarchaeon Halorubrum lacusprofundi (29). Isolates of Haloarcula spp. and Haloferax sp. obtained from a potash mine crystallization pond in north Germany had unsaturated ether lipids up to 37% of the total membrane lipid content. Only the phospholipids were unsaturated, and these contained mostly four or six double bonds in the archaeol chain.
Novel types of acylhalocapnines were described by Angela Corcelli (Bari, Italy). Salisaeta longa, an organism that requires lower salt concentrations than the related Salinibacter(Bacteroidetes), contains the hydroxyl fatty acid ester of 2-carboxy-2-amino-3,4-hydroxy-17-methyloctadec-5-ene-1-sulfonic acid, a sulfonate sphingoid base for which the common name of halocapnine is suggested (12). Salinibacter contains similar acylhalocapnine lipids in its membrane, as well as a retinal protein named xanthorhodopsin and an unusual ketocarotenoid named salinixanthin, found in a 1:1 molar ratio with the retinal pigment. Janos Lanyi (Irvine, CA) showed how the two chromophores interact and how the carotenoid acts as an antenna, transferring the absorbed light energy to the xanthorhodopsin proton pump. Such an energy transfer phenomenon appears to be unique for the clade that includes xanthorhodopsin as it is not found between the carotenoid bacterioruberin and bacteriorhodopsin in Halobacterium and related genera. The efficiency of the energy transfer is about 50%. The three-dimensional structure of the xanthorhodopsin-salinixanthin system has been determined from X-ray diffraction of xanthorhodopsin crystals, showing how the carotenoid interacts with the retinal protein (42).
The Haloquadratum walsbyi genome encodes two different bacteriorhodopsins. Both are expressed in the cells. Angela Corcelli (Bari, Italy) reported the isolation of the two forms of bacteriorhodopsin from Haloquadratum cultures using a biochemical approach. Bacteriorhodopsin was also recovered from biomass collected from the saltern crystallizer ponds of the Margarita di Savoia saltern.
Mecky Pohlschröder (Philadelphia, PA) presented recent results pertaining to mechanisms of protein transport across haloarchaeal cytoplasmic membranes. In haloarchaea, although the Sec pathway transports important substrates, including subunits of type IV pilus-like structures, the Tat pathway is used extensively and transports a wide range of secreted proteins, the majority of which appear to be anchored to the haloarchaeal membrane via a lipid anchor. In silico analyses suggest that prominent use of the Tat pathway as well as extensive anchoring of Tat substrates via a lipid anchor is unique to halophilic Archaea (the latest views on the membranal mechanisms of protein secretion in Haloferax volcanii). The Sec pathway remains an essential mode of protein transport in halophilic Archaea (80, 84). Novel programs allowing more accurate predictions of protein subcellular location (publicly available at SignalFind.org) were also presented.
DNA REPLICATION, TRANSCRIPTION, TRANSLATION, AND POSTTRANSLATIONAL MODIFICATION IN HALOPHILIC ARCHAEA
Relatively few talks dealt with the molecular biology of halophiles and the basic properties of the DNA replication, transcription, and translation machinery in different groups of halophiles.
Stuart MacNeill (St. Andrews, United Kingdom) presented novel information on the structure of the replication fork of Haloferax. Haloferax volcanii encodes a single minichromosome maintenance protein. Its N-terminal domain has a putative DNA-binding β-hairpin, a Cys4 zinc ribbon, and a β-hairpin with a role in interdomain communication. Genetic analysis of different mutants enabled the elucidation of the roles of the different proteins associated with the replication fork (43).
The biosynthesis and assembly of gas vesicles in halophilic Archaea have been used as a model for the study of transcription and other molecular processes in the Halobacteriaceae for nearly 3 decades now. Felicitas Pfeifer (Darmstadt, Germany) presented the latest information how the Halobacterium salinarum gas vesicle, when expressed in Haloferax volcanii, can be used as a convenient model system for the study of gene expression, transcription, and translation. Gas vesicles are composed of two structural proteins: the hydrophobic GvpA that forms the core of the cylinders and the hydrophilic GvpC that cross-links the GvpA subunits at the outside and provides strength to the vesicles. GvpC is now also known to be involved in the determination of the shape of the vesicles. Anaerobiosis inhibits gas vesicle formation. Fourteen gvp genes are required for gas vesicle formation, and these are arranged in two oppositely organized clusters. The function of the different promoters and transcriptional activators is becoming increasingly clear (13, 36, 81).
Updates about the molecular mechanisms of translational control in Halobacterium salinarumand Haloferax volcanii were given by Jörg Soppa (Frankfurt, Germany). Different mechanisms for translation initiation are known: (i) interaction between 16S rRNA and a Shine-Dalgarno sequence, (ii) the eukaryotic mechanism of linear scanning of the small ribosomal subunit from the 5′-cap to the start codon, (iii) an alternative eukaryotic mechanism using internal ribosomal entry sites, and (iv) leaderless transcripts that require an undissociated ribosome and the initiator tRNA (a mechanism encountered in all three domains of life). Characterization of the 5′ and 3′ ends of haloarchaeal transcripts showed that the majority of the transcripts are leaderless, that Shine-Dalgarno sequences are very rare, and that about a third do not fall in any of these four classes and must use a novel, yet uncharacterized method of translation initiation (19).
Posttranslational modification is studied in the laboratory of Jerry Eichler (Beer-Sheva, Israel), centering on the biosynthesis of the glycoproteins so abundantly found in the cell envelope of the Halobacteriaceae. Asn-modified glycoproteins are common in Archaea, and their production was studied using Haloferax volcanii as a model. A series of agl (archaeal glycosylation) genes was defined, encoding proteins involved in the assembly and attachment of a novel pentasaccharide to Asn residues of the S-layer glycoprotein. The functions of several Agl proteins are now known (1, 2, 45, 91).
GENETIC SYSTEMS FOR HALOPHILIC PROKARYOTES
Two interesting systems for genetic manipulation of halophiles were highlighted at the meeting. Already in the original species description of Haloferax volcanii the formation of intercellular bridges was noted (52). Moshe Mevarech (Tel Aviv, Israel) presented a survey of the genetic manipulation of Haloferax volcanii, developed since genetic transfer based on cell mating was first described 25 years ago (50). The mating system of Haloferax volcanii resembles eukaryotic sexual mating rather than bacterial lateral gene transfer. Large amounts of genetic material can be transferred this way (3). The successful mating of Haloferax volcanii and Haloferax mediterranei, yielding hybrid progeny, was announced.
Saskia Köcher (Frankfurt, Germany) presented a (prize-winning) poster describing the establishment of a genetic system for the manipulation of Halobacillus halophilus, based on protoplast fusion and markerless gene disruption. Cells can be transformed by protoplast transformation, resulting in integration of a nonreplicating plasmid via single homologous recombination. The method was used to generate proline biosynthesis mutants. This genetic manipulation strategy will now open the way to study many more properties of Halobacillus at the genetic level.
BIOTECHNOLOGICAL APPLICATIONS OF HALOPHILES
In comparison to other groups of extremophilic microorganisms such as the thermophiles and the alkaliphiles, the halophiles of all three domains have been relatively little exploited in biotechnological processes, with notable exceptions of β-carotene from Dunaliella, bacteriorhodopsin from Halobacterium, and ectoine from Halomonas (57). The biotechnology section of the meeting focused on production/modification techniques of compatible solutes, bioplastics, and halophilic enzymes. In addition, attention was drawn toward secondary metabolites from halophiles as well as bioremediation and biofuel production.
One success story of halophile biotechnology is the production and application of the compatible solute ectoine, currently produced at large scale by bitop AG in Germany using “bacterial milking” of Halomonas elongata. Ectoine is the active ingredient of many cosmetics and skin care products and increasingly becomes important in medicinal preparations (30). In addition, ectoine (and/or suitable derivatives) is used as a protectant for biomolecules and enhancer in molecular biology applications such as PCR and DNA microarray techniques (47, 74). Erwin Galinski (Bonn, Germany) presented a survey of the industrial production processes of ectoine and, in particular, a critical analysis of the maximal specific production rates obtainable with H. elongata as the production strain (50 mg g−1 of dry weight h−1 at 5 to 7.5% NaCl). Different strategies have been applied in attempts to increase production, including heterologous expression of the ectoine gene cluster in Escherichia coli and concomitant overexpression of genes that increase the supply of limiting precursors for ectoine biosynthesis, thus bypassing “metabolic bottlenecks“ (15, 76). Heterologous expression of the ectoine gene cluster in E. coli is at present not a suitable alternative to ectoine production in H. elongata. With respect to the hydroxylated derivative (S,S-β-hydroxyectoine) the situation is, however, different. As this compound is always produced in a mixture with ectoine in H. elongata, a costly chromatographic separation is required. By overexpressing the ectD gene (encoding ectoine hydroxylase) in E. coli, an efficient whole-cell biotransformation system for ectoine has been established in which the product (hydroxyectoine) leaked out of the cells and accumulated in the medium (E. A. Galinski, M. Stein, A. Ures, and T. Schwarz, World patent application WO 2009/059783 A1). Novel developments concern use of genetically engineered H. elongata for production of rare and thus far inaccessible compatible solutes. The potential of this approach for the development of new production processes was demonstrated using the compatible solutes mannosylglycerate (gene cluster from the thermophilic Rhodothermus marinus) and N-acetyl-glutaminyl glutamine-1-amide (gene cluster from Pseudomonas putida) as examples.
Whereas in past meetings the production of extracellular halophilic polymers with interesting rheological properties had claimed attention, the emphasis of this year's meeting (as regards polymers) has clearly been on intracellular polyesters. Production of poly-β-hydroxyalkanoates—biodegradable polymers with plastic-like properties—although not restricted to halophilic prokaryotes, was the topic of no less than four talks and a number of posters. Some halophilic or halotolerant Bacteria were shown to be excellent producers of such bioplastics. One of these is Halomonas boliviensis, as argued by Jorge Quillaguamán (Cochabamba, Bolivia), who presented strategies to optimize the biosynthesis of such bioplastics coupled with production of the high-value products ectoine and hydroxyectoine (65, 86). Archaea of the genus Haloferaxare also known as poly-β-hydroxyalkanoate producers and the biosynthetic pathway leading to their production were elucidated by Hua Xiang and colleagues (Beijing, China) (33, 41). Unfortunately none of the presenters compared the potential of halophilic producers with the current productivity of industrial strains as used, for example, by Metabolix/ADM (Cambridge, MA) for their bioplastic product Tirel.
Many alkaliphiles are halophilic as well, and many useful enzymes applied in the detergent industry (washing powders), the textile industry, and other processes were derived from bacteria growing in saline alkaline lakes. Brian Jones (Leiden, Netherlands) explained how the saline alkaline lakes in Kenya and Inner Mongolia have been a rich source of organisms and/or genes from metagenomic libraries, and some of these are already explored as starting material for the production of commercially valuable enzymes, in particular, proteases and amylases.
Halophilic enzymes (typical for Archaea and Salinibacter but also for exoenzymes of any halophile) are characterized by an excess of acidic amino acids and subsequent negative surface charge. This peculiarity allows effective competition for hydration water and enables function in solutions of low water activity, including organic solvent/water mixtures. The immediate advantages for enzyme technology are as follows: increased salt and heat tolerance, a catalytic environment which enables use of less polar educts, and potential reversal of hydrolytic reactions, all of which make them strong candidates for industrial biocatalysts.
An increasingly important industrial application of enzymes is the environmentally friendly production of stereo-specific building blocks for pharmaceuticals in “white biotechnology.” One such example, the stereo-specific production of alcohols from ketones was presented by Leanne Timpson and her colleagues Ann-Kathrin Liliensiek and Francesca Paradisi (Dublin, Ireland). Halobacterial alcohol dehydrogenases were overexpressed in Haloferax volcanii, using novel expression systems (4, 39). A number of poster presentations outlined the search for useful enzymes such as proteases, cellulases, lipases, amylases, and mannanases from halophiles, including isolates from Chinese and Iranian lakes, and a prize-winning presentation by Yasuhiro Shimane and colleagues (Saitama, Japan) on enzymes derived from haloarchaea isolated from domestic and commercial salt samples.
Nayla Munawar and Paul Engel (Dublin, Ireland) approached protein engineering of substrate specificity in a halophilic enzyme by site directed mutagenesis in the absence of a crystal structure of the enzyme. Using the crystal structure and previous mutagenesis of a mesophilic counterpart (Clostridium symbiosum glutamate dehydrogenase) as a guide, they selected corresponding residues in Halobacterium salinarum glutamate dehydrogenase (GDH) for site-directed mutagenesis and created a novel halophilic dehydrogenase which accepts l-methionine, l-norleucine, and l-norvaline as substrates instead of glutamate.
Secondary metabolites and, in particular, the untapped potential of halophilic actinomycetes as a source for novel antibiotics are increasingly becoming important, as explained by Wen-Jun Li (Kunmin, China). The abundance of culturable yet unknown types was beyond expectation, with predominance of Nocardiopsis, Saccharomonospora, and Streptomonospora. In the context of new drug discoveries, Xiukun Lin (Qingdao, China) reported on novel compounds from actinomycetes isolated from salterns and their cytotoxic effect against a range of cancer cell lines.
The worldwide problem of petroleum contamination and potential application of halophiles for bioremediation were addressed by Mohammad Amoozegar (Tehran, Iran), who described a novel isolate, similar to Alcanivorax dieselolei, able to grow on crude oil, diesel fuel, and pure aliphatic hydrocarbons but unable to degrade aromatic compounds. Its use in saline soils was investigated. A consortium of at least six culturable strains (including Marinobacter and Halomonas sp.) was able to degrade various polyaromatic hydrocarbons over a salinity range from 1 to 17% NaCl. Thus, the degrading potential of halophiles has just started to come to light and will become increasingly important in the future.
Another process in which halophiles may contribute in the future is the production of biofuel. Melanie Mormile (Rolla, MO) explained how halophilic/haloalkaliphilic and halotolerant bacteria could be used to break down biomass material and form biofuel products. Lignocellulosic biomass as a source for fermentative production of biofuel products, such as ethanol and hydrogen, may become a commercially interesting option, provided lignin components can be removed. The required alkaline pretreatment (to remove lignin) and subsequent partial neutralization will create an environment for halophilic or haloalkaliphilic fermentative bacteria in cellulose-converting processes. The general trend toward use of algae for biofuel (biodiesel) production is problematic because of the high consumption of fresh water. The use of halophilic algae may overcome such hurdles by means of efficient nonpotable water recycling and open up a bright future for halophile technology.
It is thus possible that in the future the biotechnological application of halophiles, or of genes derived from them, will extend to many more members of this extremely diverse group of microbes. Possible areas of exploitation may stretch from production of valuable compounds and remediation of contaminated waters and soils to future solutions of the world's liquid fuel crisis.

Figure 9. Growth rate vs osmolarity for different classes of procaryotes. Osmolarity is determined by solute concentration in the environment. Osmolarity is inversely related to water activity (Aw), which is more like a measure of the concentration of water (H2O) in a solution. Increased solute concentration means increased osmolarity and decreased Aw. From left to right the graph shows the growth rate of a normal (nonhalophile) such as E. coli or Pseudomonas, the growth rate of a halotolerant bacterium such as Staphylococcus aureus, and the growth rate of an extreme halophile such as the archaean Halococcus. Note that a true halophile grows best at salt concentrations where most bacteria are inhibited.
Table 11. Limiting water activities (Aw) for growth of certain procaryotes.
Osmotolerant bacteria are those bacteria that can tolerate high osmolarity. Osmolarity is established when there are solutions of different concentrations on either sides of a semipermeable membrane. High osmolarity would be described as a solution containing high concentration of solute. So basically these bacteria are capable of tolerating such high concentrations of solute in the surrounding hypertonic solution. At this solute concentration, normally other bacteria that are non-osmotolerant would die by plasmolysis due to exosmosis of water from the bacterial cell into the surrounding. But these bacteria utilize a strategy to survive in such conditions by storing compatible solutes in their cytoplasm like glycerol, glycine, betaine, sorbitol, mannitols, amino acids etc. These molecules give them the strength to survive the high osmotic stress by maintaining a cytoplasm concentration that allows influx of water into the cell.
Abstract
Halophiles, salt‐loving organisms that flourish in saline environments, are classified as slight, moderate or extreme, depending on their requirement for sodium chloride. While most marine organisms are slight halophiles, moderate and extreme halophiles are generally more specialised microbes inhabiting hypersaline environments found all over the world in arid, coastal and deep‐sea locations, underground salt mines and artificial salterns. Halophilic microorganisms include heterotrophic, phototrophic and methanogenic archaea, photosynthetic, lithotrophic and heterotrophic bacteria and photosynthetic and heterotrophic eukaryotes. Examples of extremely halophilic microorganisms include Halobacterium sp. NRC‐1, an archaeon; Aphanothece halophytica, a cyanobacterium; and Dunaliella salina, a green alga. Common multicellular halophilic eukaryotes include brine shrimp and brine fly larvae that serve as an important food source for birds. In order to balance the osmotic stress of hypersaline environments, halophilic microorganisms either accumulate organic compatible solutes internally, produce acidic proteins to increase solvation and improve function in high salinity, or use a combination of strategies.
Key Concepts
- Halophiles are salt‐loving organisms that inhabit saline and hypersaline environments and include prokaryotic (archaeal and bacterial) and eukaryotic organisms.
- Halophiles may be classified as slight, moderate or extreme, and as obligate halophiles or halotolerant.
- Many halophiles accumulate compatible solutes in cells to balance the osmotic stress in their environment.
- Some halophiles produce acidic proteins that function in high salinity by increasing solvation and prevent protein aggregation, precipitation and denaturation.
- Halophiles and their biomolecules are useful for applications in biotechnology, medicine and industry.
Keywords: archaea; Artemia; biotechnology; Dunaliella; Halobacterium; hypersaline environments; microbial diversity; microbial mat; osmotic protection; salt resistance
Introduction

Halophilic extremophiles, or simply halophiles, are a group of microorganisms that can grow and often thrive in areas of high salt (NaCl) concentration. These hypersaline areas can range from the salinity equivalent to that of the ocean (~3-5%), up to ten times that, such as in the Dead Sea (31.5% average 3). Halophiles have been found belonging to each domain of life but primarily consist of archaea.14 They are metabolically diverse, ranging from simple fermenters to iron reducers and sulfide oxidizers.
Physical Environment
Location
Naturally-Occurring Hypersaline Environments
Hypersaline environments are present on each continent and are primarily found in “arid and semi-arid regions.”17 Some are commonly known, like Utah’s Great Salt Lake, or the Dead Sea between Israel and Jordan. Others are less well known, such as Antarctica’s Deep Lake or Papua New Guinea’s undersea geothermal vents.6 19

Constructed Hypersaline Environments
Anthropogenic hypersaline environments are commonly created by the salt industry. Salterns are large ponds that are filled with saltwater from the ocean or another source that are then evaporated away. In this process, the salinity of the water gradually increases as water evaporates until it reaches saturation (~26% at 20°C). The salt then precipitates out and is harvested. Halophiles take advantage of this environment and often their presence becomes visible due to pigments they produce.
Conditions
Salt
| Halophile Classification13 | |||
|---|---|---|---|
| Slight Halophile | Moderate Halophile | Extreme Halophile | |
| Percent Salt | 2 - 5% | 5 - 20% | 20 - 30% |
| Molarity | 0.34 - 0.85 M | 0.85 - 3.4 M | 3.4 - 5.1 M |
Often in hypersaline environments, salts other than NaCl are also present. A salt profile for the Dead Sea is shown below. However, because halophiles are defined in relation to NaCl concentration, other salt content is not considered for halophilic classification.
| Salt Profile of the Dead Sea 3 | |||||||
|---|---|---|---|---|---|---|---|
| Salt Type | MgCl2 | NaCl | CaCl2 | KCl | MgBr2 | CaSO4 | RbCl |
| Percent Total Salts | 51.26% | 28.19% | 13.64% | 4.57% | 2.17% | 0.10% | 0.03% |
| Salinity and Temperature Measurements for Saline Environments.3 6 | ||||
|---|---|---|---|---|
| Pacific Ocean | Deep Lake, Antarctica | Great Salt Lake, USA | Dead Sea, Israel/Jordan | |
| NaCl Percent (g/L) | 3.4 - 3.7% | 21 - 28% | 12 - 33% | 31.5 - 34.2% |
| Temperature (C) | 1.4 - 30 C | 0 - 11.5 C | -5 - 35 C | 21 - 36 C |
Temperature
Often in hypersaline environments, the salinity is just one extreme microbes must overcome. Hypersalinity often co-occurs with extreme temperature conditions, both hot and cold.
Psychrophiles
In the case of Deep Lake, Antarctica, extreme cold and high salinity meet. Deep Lake has salinity levels ranging from 21-28%, putting the halophiles present in the extreme halophile classification. Psychrophiles are “organisms having an optimal temperature for growth at about 15°C or lower, a maximal temperature for growth at about 20°C, and a minimal temperature for growth at 0°C or below”.11 In the case of Deep Lake, eight months of the year are spent below 0°C, with a yearly maximum temperature of only 11.5°C.6 Organisms that survive and thrive here are not only halophiles, but also psychrophiles. Similar overlaps of cold and hypersaline environments have been found in other places as well.

Thermophiles
In the case of the black smokers off the coast of Papua New Guinea, a hypersaline environment was created by a hydrothermal vent. This hydrothermal vent releases a hypersaline effluent that was measured at greater than 250°C. Samples taken from the internal walls of the smoker contained Halomona and Haloarcula species.19 This classifies Halomona and Haloarcula as both thermophiles and halophiles. Such overlapping conditions of hypersalinity and extreme heat are also present in some hot springs and other hydrothermal vents.
Other Extremes
Some hypersaline environments have been found that overlap with other extremes, such as low and high pH, and dry, desiccating conditions. Organisms in such conditions would be considered haloacidophiles, haloalkaliphiles, and haloxerophiles, respectively.
Microbial Communities
Archaea
The majority of extreme halophiles are archaea 14. A thorough study of salt-crystallizing ponds from several places around the world by Oren in 2002 showed consistent communities between saltern ponds. The genera isolated were primarily Haloferax, Halogeometricum, Halococcus, Haloterrigena, Halorubrum, Haloarcula, Halobacterium, and Natronococcus. At lower degrees of salinity, diversity of genera present increases.
Bacteria
Bacteria belonging to the genus Salinibacter have been found to be “no less salt-dependent and salt-tolerant than the most halophilic among the Archaea…” 14 As such, within specific extreme environments, some bacteria “may coexist with the halophilic archaeal community .”14 As with archaea, bacterial diversity also increases as salinity decreases.
Eukarya
At the extreme end of hypersalinity, eukarya are absent. However, there are a few moderately or slightly halophilic eukaryotes that can contribute to the halophilic communities. There are also types of halophilic fungi, such as Debaryomyces hansenii and Hortaea werneckii .47
Microbial Processes
Metabolism
According to Oren in 2002, “The metabolic diversity of halophiles is great as well: they include oxygenic and anoxygenic phototrophs, aerobic heterotrophs, fermenters, denitrifiers, sulfate reducers, and methanogens.” Additionally, halophilic sulfide oxidizers, iron-reducers, and acetogens have also been discovered.202123 Halophilic bacteria have been found to perform fermentation, acetogenesis, sulfate reduction, phototrophy, and methanogenesis.13
Mechanisms for Saline Resistance
Compatible Solutes
One mechanism halophiles use to survive in high concentrations of salt is the synthesis of osmoprotectants, which are also known as compatible solutes. These work by balancing the internal osmotic pressure with the external osmotic pressure, making the two solutions isotonic, or close to it. Compatible solutes are small-molecular weight molecules. “Some are widespread in microorganisms, namely trehalose, glycine betaine and α-glutamate, while others are restricted to a few organisms.”16 The use of compatible solutes is the most common mechanism of salt resistance among halophiles.
Potassium Control
A second, less common mechanism of defense against salt is through controlling potassium levels. This mechanism is performed by pumping in large amounts of K+ ions into the cytoplasm. To deal with the increased solute level, and to increase the solubility of the proteins within the cytoplasm, many proteins have evolved to become more acidic.16
Current Research
Diversity and Systematics Research
A large number of research projects have been focused on finding and classifying novel halophiles.
A paper by Ndwigah in 2016 isolated a halophilic fungus, determining its optimal growth conditions and its metabolites.12
McDuff in 2016 characterized several CO-oxidizing halophiles. This paper also provided a novel species description of Haloferax namakaokahaiae.10
Amoozegar et al. provided a novel species description of a halophilic bacterium, Oceanobacillus halophilus. This bacterium was isolated from a brine lake in Iran.1
A paper by Ndwigah in 2016 isolated a halophilic fungus, determining its optimal growth conditions and its metabolites.12
McDuff in 2016 characterized several CO-oxidizing halophiles. This paper also provided a novel species description of Haloferax namakaokahaiae.10
Amoozegar et al. provided a novel species description of a halophilic bacterium, Oceanobacillus halophilus. This bacterium was isolated from a brine lake in Iran.1
Protein Research
Because of the unique conditions to which the proteins of halophiles are subjected, their proteins are heavily studied, especially for their use in industry.
Varun in 2009 looked at the possibility of using a specific halophile to generate electricity. This species under its normal growth conditions fermented glycerol to ethanol, and produced electricity through iron reduction. According to the paper, “Glycerol is a major byproduct of biodiesel industry and therefore bacterial fermentation of this glycerol to ethanol would help to manage the waste as well as produce a value-added product.” This species could be potentially used in microbial fuel cells.21
A paper by Calegari-Santos in 2016 also looked at using halophiles in industry to produce carotenoids. These carotenoids have numerous “potential uses in Biotechnology and Biomedicine.”5
Varun in 2009 looked at the possibility of using a specific halophile to generate electricity. This species under its normal growth conditions fermented glycerol to ethanol, and produced electricity through iron reduction. According to the paper, “Glycerol is a major byproduct of biodiesel industry and therefore bacterial fermentation of this glycerol to ethanol would help to manage the waste as well as produce a value-added product.” This species could be potentially used in microbial fuel cells.21
A paper by Calegari-Santos in 2016 also looked at using halophiles in industry to produce carotenoids. These carotenoids have numerous “potential uses in Biotechnology and Biomedicine.”5
Halovirus Research
The ubiquity of viruses does not stop when the environment gets salty. Viral infection in hypersaline environments is also being actively investigated.
Atanasova in 2016 investigated how haloviruses and their hosts interact on an ecology level, and gave general lifestyle descriptions of numerous haloviruses. 2
Pietila in 2016 worked on the classification of haloviruses, proposing a new family for “archaeal pleomorphic viruses with single-stranded or double-stranded DNA genomes.”
Atanasova in 2016 investigated how haloviruses and their hosts interact on an ecology level, and gave general lifestyle descriptions of numerous haloviruses. 2
Pietila in 2016 worked on the classification of haloviruses, proposing a new family for “archaeal pleomorphic viruses with single-stranded or double-stranded DNA genomes.”
Asked by: Dave Cullis, Leicester
Salt kills some types of bacteria, effectively by sucking water out of them. In a process known as osmosis, water passes out of a bacterium so as to balance salt concentrations on each side of its cell membrane. Without water, bacterial proteins such as enzymes cannot function and eventually the cell collapses in on itself.
Some bacteria can tolerate salt; they are halotolerant. Certain strains of Staphylococcus, responsible for infections, blood poisoning, and even death, are halotolerant. These pathogens have a salt alert system that uses sponge-like molecules to prevent water loss.
Archaebacteria är˝kēbăktĭr´ēə [key], diverse group of bacteria (prokaryotes), sometimes called the archaea and considered a major group unto themselves. Archaebacteria are contrasted with the Eubacteria, from which they differ biochemically in the arrangement of the bases in their ribosomal RNA and in the composition of their plasma membranes and cell walls. There are three major known groups of Archaebacteria: methanogens, halophiles, and thermophiles. The methanogens are anaerobic bacteria that produce methane. They are found in sewage treatment plants, bogs, and the intestinal tracts of ruminants. Ancient methanogens are the source of natural gas. Halophiles are bacteria that thrive in high salt concentrations such as those found in salt lakes or pools of sea water. Thermophiles are the heat-loving bacteria found near hydrothermal vents and hot springs. Many thermophiles are chemosynthetic (seechemosynthesis ), using dissolved sulfur or other elements as their energy source and iron as a means of respiration. Archaebacteria emerged at least 3.5 billion years ago and live in environments that resemble conditions existing when the earth was young.
Living environment[edit]

Haloarchaea require salt concentrations in excess of 2 M (or about 10%) to grow, and optimal growth usually occurs at much higher concentrations, typically 20–25%. However, Haloarchaea can grow up to saturation (about 37% salts).[17]
Haloarchaea are found mainly in hypersaline lakes and solar salterns. Their high densities in the water often lead to pink or red colourations of the water (the cells possessing high levels of carotenoid pigments, presumably for UV protection).[18] Some of them live in underground rock salt deposits, including one from middle-late Eocene (38-41 million years ago).[19] Some even older ones from more than 250 million years ago have been reported.[20]
Adaptations to environment[edit]
Haloarchaea can grow at an aw close to 0.75, yet a water activity (aw) lower than 0.90 is inhibitory to most microbes.[21] The number of solutes causes osmotic stress on microbes, which can cause cell lysis, unfolding of proteins and inactivation of enzymes when there is a large enough imbalance.[22] Haloarchaea combat this by retaining compatible solutes such as potassium chloride (KCl) in their intracellular space to allow them to balance osmotic pressure.[23] Retaining these salts is referred to as the “salt-in” method where the cell accumulates a high internal concentration of potassium.[24] Because of the elevated potassium levels, haloarchaea have specialized proteins that have a highly negative surface charge to tolerate high potassium concentrations.[25]
Haloarchaea have adapted to use glycerol as a carbon and energy source in catabolic processes, which is often present in high salt environments due to Dunaliella species that produce glycerol in large quantities.[24]
Phototrophy[edit]
Bacteriorhodopsin is used to absorb light, which provides energy to transport protons (H+) across the cellular membrane. The concentration gradient generated from this process can then be used to synthesize ATP. Many haloarchaea also possess related pigments, including halorhodopsin, which pumps chloride ions in the cell in response to photons, creating a voltage gradient and assisting in the production of energy from light. The process is unrelated to other forms of photosynthesis involving electron transport, however, and haloarchaea are incapable of fixing carbon from carbon dioxide.[26] Early evolution of retinal proteins has been proposed as the purple Earth hypothesis.[5]
Cellular shapes[edit]
Haloarchaea are often considered pleomorphic, or able to take on a range of shapes—even within a single species. This makes identification by microscopic means difficult, and it is now more common to use gene sequencing techniques for identification instead.
One of the more unusually shaped Haloarchaea is the "Square Haloarchaeon of Walsby". It was classified in 2004 using a very low nutrition solution to allow growth along with a high salt concentration, square in shape and extremely thin (like a postage stamp). This shape is probably only permitted by the high osmolarity of the water, permitting cell shapes that would be difficult, if not impossible, under other conditions.
As exophiles[edit]
Haloarchaea have been proposed as a kind of life that could live on Mars; since the Martian atmosphere has a pressure below the triple point of water, freshwater species would have no habitat on the Martian surface. The presence of high salt concentrations in water lowers its freezing point, in theory allowing for halophiles to exist in saltwater on Mars.[27]
Haloarchaea (halophilic archaea, halophilic archaebacteria, halobacteria)[1] are a class of the Euryarchaeota,[2] found in water saturated or nearly saturated with salt. Halobacteria are now recognized as archaea, rather than bacteria and are one of the largest groups. The name 'halobacteria' was assigned to this group of organisms before the existence of the domain Archaea was realized, and while valid according to taxonomic rules should be updated.[3] Halophilic archaea are generally referred to as haloarchaea to distinguish them from halophilic bacteria.
These microorganisms are members of the halophile community, in that they require high salt concentrations to grow, with most species requiring more than 2.0M NaCl for growth and survival.[4] They are a distinct evolutionary branch of the Archaea distinguished by possession of ether-linked lipids and the absence of murein in their cell walls.
Haloarchaea can grow aerobically or anaerobically. Parts of the membranes of haloarchaea are purplish in color,[5] and large blooms of haloarchaea appear reddish, from the pigment bacteriorhodopsin, related to the retinal pigment rhodopsin, which it uses to transform light energy into chemical energy by a process unrelated to chlorophyll-based photosynthesis.
Haloarchaea have a potential to solubilize phosphorus. Phosphorus-solubilizing halophilic archaea may well play a role in P (phosphorus) nutrition to vegetation growing in hypersaline soils. Haloarchaea may also have application as inoculants for crops growing in hypersaline regions.[6]
Osmotic Pressure and Water Availability:
Osmotic pressure is the force water exerts on the semi-permeable membrane (plasma membrane) surrounding the cell. Water moves across the plasma membrane in response to an unequal distribution of dissolved solutes in the environment. When a microorganism is placed into a hypotonicenvironment/medium in which the concentration of solute is lower outside the cell than inside the cell, water flows into the cell. If this influx of water is uncontrolled, the cell may eventually burst. Most bacteria, algae and fungi have rigid cell walls that allow them to tolerate and even enjoy a somewhat hypotonic environment. Many microorganisms keep the solute concentration slightly higher in their protoplasm by synthesizing solutes that are compatible with their metabolism and growth, thus keeping their membrane pushed tightly against the cell wall. By contrast, when microorganisms are placed into a hypertonicenvironment where the concentration of solute is higher outside the cell, water is lost from the cell resulting in dehydration, shrinkage of the plasma membrane and eventual death. This is a process known as plasmolysis.
As can be seen, the osmotic concentration of the environment affects the water that is available to a microorganism. The greater the solute concentration of the environment, the less readily available the water. Some prokaryotes can maintain the availability of water in environments with high solute concentrations (hypertonic environments) by increasing the solute concentration within the cell. Microorganisms that can do this and thus tolerate hypertonic environments are osmotolerant. Osmotolerant bacteria, such as Staphylococcus aureus can grow in a wide variety of environments with varying osmotic pressures. In fact this bacteria can be cultured in media containing sodium chloride (NaCl) concentrations as high as 3M.
Some bacteria specifically require an environment with a high concentration of sodium chloride. These organisms are called halophiles.
Halophiles are organisms that need salt in their environment to live. Halophiles live in evaporation ponds or salt lakes such as Great Salt Lake, Owens Lake, or Dead Sea.[1] The name "halophile" comes from Greek for "salt-loving".[2]
Most halophiles are archaeans, but some bacteria and eukaryotes are also halophiles, such as the alga Dunaliella salina.[3
Lifestyle[change | change source]
Most halophilic and salt-eating animals use energy to remove salt from their cytoplasm.[4] Normally, organisms living around a lot of salt would lose water and die because of osmosis.[1][5][6][7] Water inside the organism would move from inside the cell to its outside environment. This is because there is always movement of water to reach a state where the concentrations of salt are the same on both sides of the cell membrane.

To get to this state, halophiles use two different methods. In the first (mainly used by bacteria, some archaea, yeasts, algae and fungi), organic compounds are stored in the cytoplasm. These compounds help the organism survive the stress of osmosis.[9][10] The most commonly used solutes for this process are neutral and include amino acids and sugars.[7][11][12] Advantages to this method are that organisms can live in a wider range of salt concentrations. Also, if proteins are not exposed to high salt levels, they do not have to adapt to high salt levels. It requires the organism to use much more energy then the adaption below.[4]
The second, less common adaptation, is the selective intake of potassium (K+) ions into the cytoplasm. In exchange, the organism pumps sodium (Na+) ions out with the help of the sodium-potassium pump.[12] Ions of sodium may be used instead of potassium, but potassium is the most common.[7] This adaptation is only used by one order of bacteria and one family of Archaea.[13] An advantage to this method is that it uses much less energy than the adaption above. The main disadvantage to this adaption is that all of the machinery inside the cell (enzymes, structural proteins, etc.) must be adapted to high levels of non-organic ions,[13] and high salt levels.[4] This is much more demanding than the adaptation described above.
Most halophilic organisms only use one of the two methods, but a few halophiles can use both.[14]
Categorization[change | change source]
Halophiles are categorized by the levels of salt at which they grow best. Halophiles can either be categorized as slight halophiles, moderate halophiles, or extreme halophiles.[15]Slight halophiles grow best in concentrations of salt around 2% to 5%. An example of a slight halophile is Erythrobacter flavus.[16] Slight halophiles live in mud on the ocean floor, in seawater, and in garden soil.[17]
Moderate halophiles grow best in concentrations of salt around 5% to 20%. Examples of moderate halophiles are organisms in the genera Desulfovibrio, Desulfocella, Desulfohalobium, and Desulfotomaculum. Moderate halophiles live in marine salterns, saline lakes, in matter at the bottom of the sea, and in oil brine fields.[18]
Extreme halophiles grow best in concentrations of salt from 20% to 30%. Examples of extreme halophiles are Salinibacter ruber and organisms in the Halobacteria class. Extreme halophiles live in the Dead Sea in the Middle East and in man-made solar salterns (lakes used for making sea salt).
Importance and uses[change | change source]
In nature[change | change source]

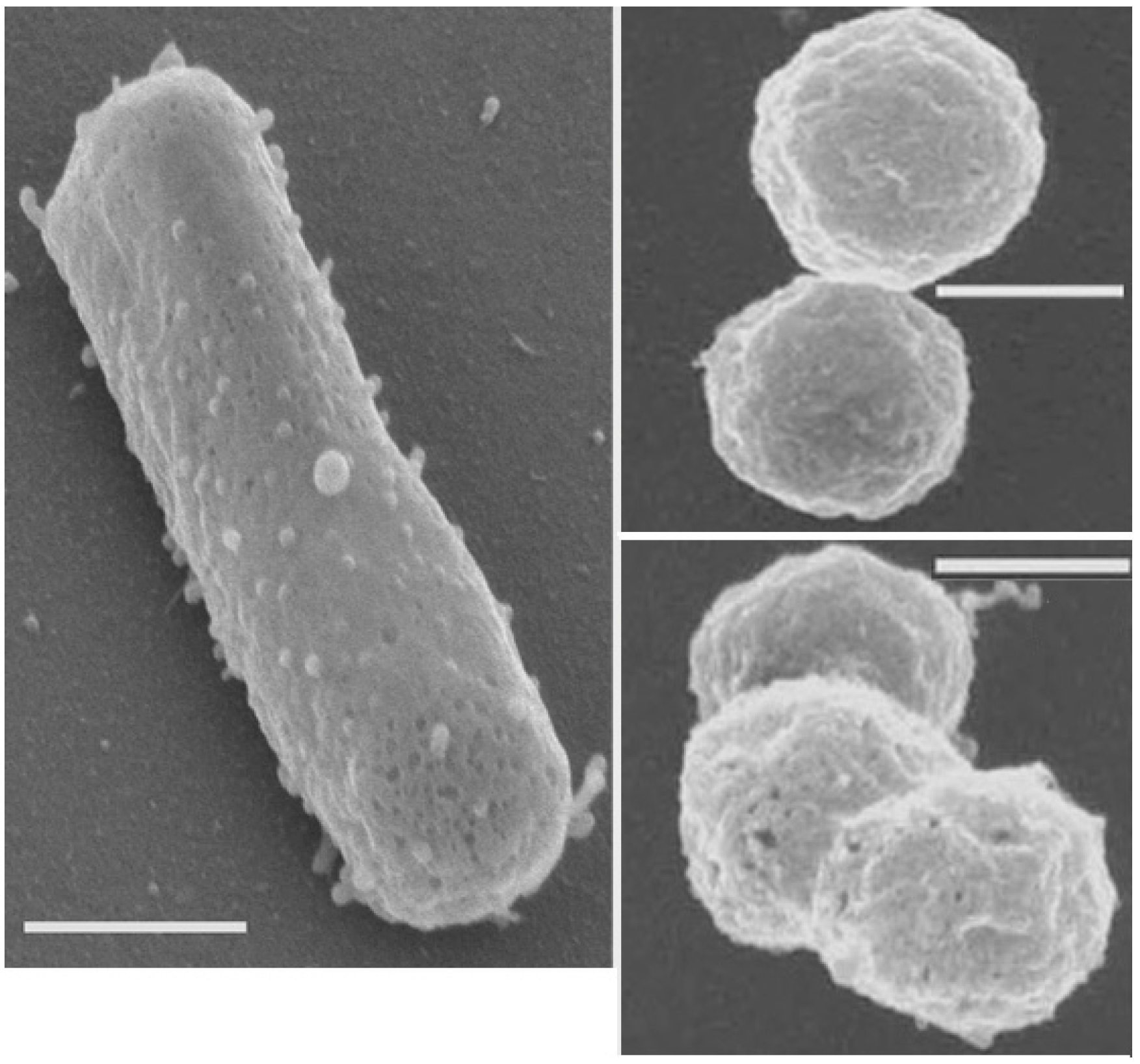{kind=link}
{kind=link}
{kind=link}
Halophiles play an important part in ecosystems. For example, halophiles often support entire populations of wild birds.
Halophiles are useful for cleaning up polluted environments.[19] Waste water with salt concentrations more than 2% is ideal for halophiles to remove organic pollutants from.[20] For instance, halophiles have been shown to remove phenol (a poisonous chemical) from their environments. This could lead to future use in cleaning up oil spills.[21]
In fermentation[change | change source]
Halophiles play an important part in the fermentation of some foods. For instance, halophiles ferment soy and fish sauces.[22] Halophiles also ferment salted fish.
In biotechnology[change | change source]
Halophilic microorganisms are useful in biotechnology. The compounds that certain halophiles make are valuable. Some of these compounds are not found anywhere else in the living world.[23] The salt-tolerant enzymes that halophiles produce can be used in a variety of ways.[24] For example, these enzymes could be used for rough industrial processes, like food processing.[19]
Some moderate halophiles produce sugars outside the cell. These sugars can be used as thickeners and emulsifiers in the petroleum and medicine industries.[25
Water Availability
Water is the solvent in which the molecules of life are dissolved, and the availability of water is therefore a critical factor that affects the growth of all cells. The availability of water for a cell depends upon its presence in the atmosphere (relative humidity) or its presence in solution or a substance (water activity). The water activity (Aw) of pure H2O is 1.0 (100% water). Water activity is affected by the presence of solutes such as salts or sugars, that are dissolved in the water. The higher the solute concentration of a substance, the lower is the water activity and vice-versa. Microorganisms live over a range of Aw from 1.0 to 0.7. The Aw of human blood is 0.99; seawater = 0.98; maple syrup = 0.90; Great Salt Lake = 0.75. Water activities in agricultural soils range between 0.9 and 1.0.
The only common solute in nature that occurs over a wide concentration range is salt [NaCl], and some microorganisms are named based on their growth response to salt. Microorganisms that require some NaCl for growth are halophiles. Mild halophiles require 1-6% salt, moderate halophiles require 6-15% salt; extreme halophiles that require 15-30% NaCl for growth are found among the archaea. Bacteria that are able to grow at moderate salt concentrations, even though they grow best in the absence of NaCl, are called halotolerant. Although halophiles are "osmophiles" (and halotolerant organisms are "osmotolerant") the term osmophilesis usually reserved for organisms that are able to live in environments high in sugar. Organisms which live in dry environments (made dry by lack of water) are called xerophiles.
The concept of lowering water activity in order to prevent bacterial growth is the basis for preservation of foods by drying (in sunlight or by evaporation) or by addition of high concentrations of salt or sugar.
Figure 9. Growth rate vs osmolarity for different classes of procaryotes. Osmolarity is determined by solute concentration in the environment. Osmolarity is inversely related to water activity (Aw), which is more like a measure of the concentration of water (H2O) in a solution. Increased solute concentration means increased osmolarity and decreased Aw. From left to right the graph shows the growth rate of a normal (nonhalophile) such as E. coli or Pseudomonas, the growth rate of a halotolerant bacterium such as Staphylococcus aureus, and the growth rate of an extreme halophile such as the archaean Halococcus. Note that a true halophile grows best at salt concentrations where most bacteria are inhibited.
Table 11. Limiting water activities (Aw) for growth of certain procaryotes.
| Organism | Minimum Aw for growth |
| Caulobacter | 1.00 |
| Spirillum | 1.00 |
| Pseudomonas | .91 |
| Salmonella/E. coli | .91 |
| Lactobacillus | .90 |
| Bacillus | .90 |
| Staphylococcus | .85 |
| Halococcus | .75 |
Osmotolerant bacteria are those bacteria that can tolerate high osmolarity. Osmolarity is established when there are solutions of different concentrations on either sides of a semipermeable membrane. High osmolarity would be described as a solution containing high concentration of solute. So basically these bacteria are capable of tolerating such high concentrations of solute in the surrounding hypertonic solution. At this solute concentration, normally other bacteria that are non-osmotolerant would die by plasmolysis due to exosmosis of water from the bacterial cell into the surrounding. But these bacteria utilize a strategy to survive in such conditions by storing compatible solutes in their cytoplasm like glycerol, glycine, betaine, sorbitol, mannitols, amino acids etc. These molecules give them the strength to survive the high osmotic stress by maintaining a cytoplasm concentration that allows influx of water into the cell.